Show expression patterns that are correlated with PDGFR signaling, a pattern with prominent expression of OLIG2 and other genes involved in CNS development referred to as “proneural”. PDGF ligands are upregulated in at least a third of surgical glioma samples and human glioma cell lines. The importance of PDGF signaling is underscored in genetically engineered rodent gliomas, where overproduction of human PDGFb ligand is sufficient to induce gliomagenesis in a dosedependent manner and allows to recapitulate the histologic, etiologic and pathobiologic character of the PDGF subset of human gliomas. Additionally, infusion of PDGF into the ventricles induces proliferation of the SVZ, resulting in lesions with some characteristics of gliomas. Similar to human gliomas, mouse gliomas are cellularly and molecularly heterogeneous. Glioma progression in humans is associated with deletion of the INK4A/ARF locus and loss of PTEN expression resulting in activation of Akt. The standard view of gliomagenesis is that sequential mutations occur and accumulate in cells derived from the glioma cell-of-origin. Indeed, many surgical GBM samples in patients appear clonal, with all tumor cells seemingly derived from the same cell; however, this may not necessarily mean they are derived from the cell-of-origin. Cellular heterogeneity and reports of human gliomas comprised of several genetically unrelated clones suggest the possibility of oncogenic transformation in cells not derived from the glioma cell-of-origin. The interconversion between human glioma subtypes upon recurrence and the existence of recurrent gliomas that lack mutations or deletions found in the original tumor further indicate the possibility for an expansion of an aggressive clone not arising from the cell-of-origin. In fact, PDGF-induced gliomas arising in both adult and neonatal rats have been shown to contain normal stem and progenitor cells “recruited” into glioma mass and induced to proliferate, indicating that proliferative stem-like portions of the tumor can arise from normal progenitors. However, the Mepiroxol precise nature and specific functional characteristics of these “recruited” stem or progenitor cells have not been described. Genetic analysis of surgical samples of human gliomas merely provides retrospective static information with regards to tumor evolution; lineage tracing from the cell-of-origin cannot be done in humans. Moreover, identifying and distinguishing GBM cells from the surrounding stroma is not a trivial task – glioma cells are often defined histologically, demonstrating high mitotic indices, expression of stem or progenitor cell markers, abnormal global gene expression patterns, presence of genetic alterations, and the ability to serially transplant the disease. To investigate cellular contributions and structural/functional characteristics of “recruited” cells in murine gliomas during tumor progression, we used RCAS/tv-a and the Gomisin-D bacTRAP systems. Defining tumor cells by histologic criteria, genetic analysis, global gene expression profiling and transplantation studies, we studied the clonality of mouse gliomas with respect to the cell-of-origin. Here we show that in murine gliomas induced by human PDGFb, glioma progression can occur 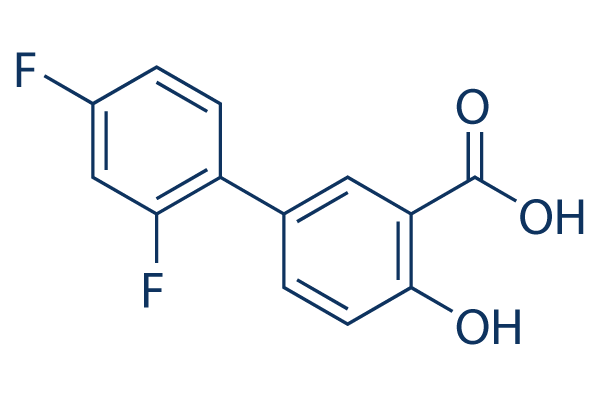 by expansion of the recruited cells, and that these cells unrelated to glioma cell-of-origin can be corrupted to become bona fide tumor. It has been recently shown that gliomas induced in adult or neonatal rats by hPDGFb-expressing retroviruses contain stem or progenitor-like cells expressing neural markers, that are contributing to glioma mass and are induced to proliferate by glioma environment.
by expansion of the recruited cells, and that these cells unrelated to glioma cell-of-origin can be corrupted to become bona fide tumor. It has been recently shown that gliomas induced in adult or neonatal rats by hPDGFb-expressing retroviruses contain stem or progenitor-like cells expressing neural markers, that are contributing to glioma mass and are induced to proliferate by glioma environment.