Therefore, the parasite must have strategies to cope with the challenges presented by such environmental variations as well as mechanisms to adapt to and take advantage of them. In this sense, the intestinal environment of the insect host is particularly relevant, and the microbes and chemicals encountered in such an environment are likely to affect the parasite-host interaction and the fate of the association. Indeed, many molecules present in the intestinal tract of triatomine hosts have been shown to affect T. cruzi. For instance, the hemolytic factor present in the stomach of R. prolixus has the capacity to lyse the epimastigote forms of some 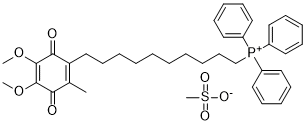 T. cruzi strains but not others. Lectins can agglutinate some T. cruzi strains. Additionally, studies of bacterial strains isolated from the triatomine gut revealed that strains of Serratia marcescens, the predominant bacterial species found in Rhodnius and Triatoma, are able to lyse epimastigote forms of T. cruzi. Altogether, these observations show that a myriad of compounds found in the triatomine gut have a decisive influence in the infectious process and determine whether T. cruzi can successfully establish itself in the vector host, which will subsequently determine its capacity to spread through human populations. We have recently shown that although the intestinal microbiota of triatomines is lacking in bacterial species diversity, it displays marked specificity with certain bacteria associated with different insect vectors. Understanding the molecular basis of the vector-parasite-commensal bacteria triad may aid in our knowledge of their determinants and the epidemiology of T. cruzi nfections in humans and in the identification of promising Lomitapide Mesylate therapeutics. Therefore, we set out to study the chemical environment of the triatomine gut through a powerful, highthroughput metabolomics approach. Our results indicate that metabolic variation among triatomine species is obvious even in laboratory conditions. We describe the identification of uniform and variable cores of metabolites in three different triatomine species. Our results provide a basis for further investigations of the interplay between T. cruzi and the triatomine digestive tract and how this may come into play during vector colonization, especially in the hindgut, where parasite metacyclogenesis occurs. Ultimately, understanding metabolic variations may allow the identification of factors regulating parasite growth and assist in the development of anti-parasitic drugs. DI-FT-ICR-MS is a technique that enables the precise quantitative characterization of metabolites in a biological sample by data accumulation, allowing the fast and accurate derivation of the metabolic profile associated with a sample. However, one drawback to this method is that the qualitative characterization of each molecular specimen relies on the comparison of its molecular weight with preexisting information stored in a database. As is common with such approaches, a lack of information in databases precludes the full characterization of a set of samples. In this report, we studied the fecal metabolome of three species of triatomines, the insect vectors of T. cruzi and, hence, Chagas disease. To Chlorhexidine hydrochloride obtain putative identities of the metabolic features detected, we searched the Human Metabolome Database due to the lack of an appropriate database for triatomine samples. The HMDB has wide metabolic coverage, currently containing information on more than 40,000 metabolites, which prompted us to use this database for our studies.
T. cruzi strains but not others. Lectins can agglutinate some T. cruzi strains. Additionally, studies of bacterial strains isolated from the triatomine gut revealed that strains of Serratia marcescens, the predominant bacterial species found in Rhodnius and Triatoma, are able to lyse epimastigote forms of T. cruzi. Altogether, these observations show that a myriad of compounds found in the triatomine gut have a decisive influence in the infectious process and determine whether T. cruzi can successfully establish itself in the vector host, which will subsequently determine its capacity to spread through human populations. We have recently shown that although the intestinal microbiota of triatomines is lacking in bacterial species diversity, it displays marked specificity with certain bacteria associated with different insect vectors. Understanding the molecular basis of the vector-parasite-commensal bacteria triad may aid in our knowledge of their determinants and the epidemiology of T. cruzi nfections in humans and in the identification of promising Lomitapide Mesylate therapeutics. Therefore, we set out to study the chemical environment of the triatomine gut through a powerful, highthroughput metabolomics approach. Our results indicate that metabolic variation among triatomine species is obvious even in laboratory conditions. We describe the identification of uniform and variable cores of metabolites in three different triatomine species. Our results provide a basis for further investigations of the interplay between T. cruzi and the triatomine digestive tract and how this may come into play during vector colonization, especially in the hindgut, where parasite metacyclogenesis occurs. Ultimately, understanding metabolic variations may allow the identification of factors regulating parasite growth and assist in the development of anti-parasitic drugs. DI-FT-ICR-MS is a technique that enables the precise quantitative characterization of metabolites in a biological sample by data accumulation, allowing the fast and accurate derivation of the metabolic profile associated with a sample. However, one drawback to this method is that the qualitative characterization of each molecular specimen relies on the comparison of its molecular weight with preexisting information stored in a database. As is common with such approaches, a lack of information in databases precludes the full characterization of a set of samples. In this report, we studied the fecal metabolome of three species of triatomines, the insect vectors of T. cruzi and, hence, Chagas disease. To Chlorhexidine hydrochloride obtain putative identities of the metabolic features detected, we searched the Human Metabolome Database due to the lack of an appropriate database for triatomine samples. The HMDB has wide metabolic coverage, currently containing information on more than 40,000 metabolites, which prompted us to use this database for our studies.
Month: May 2019
Therefore the currently low number of publically available MS raw data sets despite multiple published
Within the lesions of MOG-induced EAE and TMEV-IDD. The pathogenic role of immunoglobulins in EAE remains controversial. However, several types of EAE in mice, rats, and monkeys are accompanied by B cell responses and immunoglobulin deposition. In detail, deposition of immunoglobulin has been described in MOG-induced EAE in Dark Agouti rats, one of the models used in the present meta-analysis. However, studies in B cell deficient mice have shown that B cells are not critical for the development of MOG-induced murine EAE. Experimental models with a marked oligodendrocyte dystrophy such as cuprizone-induced toxicity analogous to the MS patterns III and IV are characterized by an early-onset and marked down-regulation of genes involved in myelination. The magnitude of the transcriptional change roughly parallels the degree of demyelination, which is estimated to affect slightly more than one half of the spinal cord in MOGinduced EAE in female Dark Agouti rats. Similarly, in TMEV-IDD a down-regulation of PLP and MBP mRNA to approximately 58% of control levels has been described using insitu hybridization, and a down-regulation of MBP mRNA to approximately 70% of control levels has been described using RTqPCR. Based on the abundant apoptotic oligodendroglial cell death in pattern III, and non-apoptotic 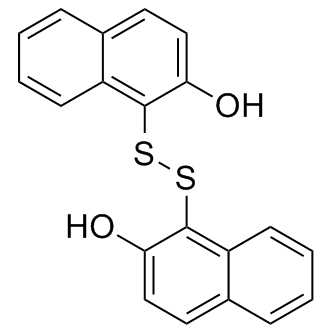 oligodendroglial cell death in pattern IV a respective differential expression of the genes of the GO term “positive regulation of apoptotic process” was suggested to differentiate between analogous conditions. Although the TNFtg model is reported to display primary oligodendrocyte apoptosis with subsequent myelin loss as the predominant pathological feature, we were unable to detect (R)-(-)-Modafinic acid corresponding transcriptional changes in the genes comprised by the selected GO terms. The 3,4,5-Trimethoxyphenylacetic acid reason for this lack of concordance remains unclear. The only GO term observed in our re-analysis of this model which is possibly linked to programmed cell death was “ion homeostasis”. It is known that TNF potently increases intracellular Ca2+ and may thereby induce apoptosis. Similar to our results, the original study describing the TNFtg data set also lacks the description of transcriptional changes associated with myelination and apoptosis. It cannot be excluded that previous studies may have overestimated the relative contribution of oligodendrocyte apoptosis in comparison to the pro-inflammatory effects of TNF-overexpression in the pathogenesis of the TNFtg model. Alternatively, it seems plausible that the induction of apoptosis in oligodendrocytes via the p55TNF receptor-signaling pathway may simply occur by activation of the preformed apoptotic cascade without any related transcriptional changes. Further studies are needed to unravel, whether alternate gene signatures are better suited or if transcriptional profiling is generally a poor approach to reflect the degree of demyelination and apoptosis. The up-regulation of apoptosis�Crelated genes in EAE is suggested to be linked to the marked infiltration of T cells, which are known for their marked expression of extrinsic apoptosis pathway genes like Fas and Fas-L. Apoptosis is known to mainly occur in lymphocytes and not in oligodendrocytes in EAE. The contribution of virus-induced oligodendrocyte apoptosis during the pathogenesis of TMEV-IDD is debated controversially, but generally regarded to be low for the TMEV BeAn-strain used in this study. Conclusively, based on the provided results, MOG- and PLP-induced EAE as well as TMEV-IDD are suggested to mimic especially MS pattern II. In contrast, the TNFtg model displayed transcriptional changes anticipated to occur in MS pattern I. The achievable level of novel information in a meta-analysis strongly depends on the availability and quality of raw data.
oligodendroglial cell death in pattern IV a respective differential expression of the genes of the GO term “positive regulation of apoptotic process” was suggested to differentiate between analogous conditions. Although the TNFtg model is reported to display primary oligodendrocyte apoptosis with subsequent myelin loss as the predominant pathological feature, we were unable to detect (R)-(-)-Modafinic acid corresponding transcriptional changes in the genes comprised by the selected GO terms. The 3,4,5-Trimethoxyphenylacetic acid reason for this lack of concordance remains unclear. The only GO term observed in our re-analysis of this model which is possibly linked to programmed cell death was “ion homeostasis”. It is known that TNF potently increases intracellular Ca2+ and may thereby induce apoptosis. Similar to our results, the original study describing the TNFtg data set also lacks the description of transcriptional changes associated with myelination and apoptosis. It cannot be excluded that previous studies may have overestimated the relative contribution of oligodendrocyte apoptosis in comparison to the pro-inflammatory effects of TNF-overexpression in the pathogenesis of the TNFtg model. Alternatively, it seems plausible that the induction of apoptosis in oligodendrocytes via the p55TNF receptor-signaling pathway may simply occur by activation of the preformed apoptotic cascade without any related transcriptional changes. Further studies are needed to unravel, whether alternate gene signatures are better suited or if transcriptional profiling is generally a poor approach to reflect the degree of demyelination and apoptosis. The up-regulation of apoptosis�Crelated genes in EAE is suggested to be linked to the marked infiltration of T cells, which are known for their marked expression of extrinsic apoptosis pathway genes like Fas and Fas-L. Apoptosis is known to mainly occur in lymphocytes and not in oligodendrocytes in EAE. The contribution of virus-induced oligodendrocyte apoptosis during the pathogenesis of TMEV-IDD is debated controversially, but generally regarded to be low for the TMEV BeAn-strain used in this study. Conclusively, based on the provided results, MOG- and PLP-induced EAE as well as TMEV-IDD are suggested to mimic especially MS pattern II. In contrast, the TNFtg model displayed transcriptional changes anticipated to occur in MS pattern I. The achievable level of novel information in a meta-analysis strongly depends on the availability and quality of raw data.
The Nupr1 gene was first described as chemical medium whose metabolite constituents are likely to affect development and infectivity
In addition to the potential Tulathromycin B effects of the uniform core of metabolites as the primary determinants of the success of the host-parasite relationship between triatomines and T. cruzi, variable metabolites may act as secondary determinants and are likely critical to the course of the host-parasite relationship. These secondary determinants may play their roles through direct actions as well as synergistic or antagonistic effects of multiple metabolites whose proportions may vary according to the insect host. It is well known that different T. cruzi strains do not have the same developmental success in different triatomine species and different ecological conditions; T. cruzi virulence varies according to the geographic location. All three species tested here are good vectors for T. cruzi. Thus, comparing the metabolome outlined here to that of triatomines with poor T. cruzi transmission efficiency would most likely aid in identifying factors involved in the development of T. cruzi. Interestingly, compounds like eicosanoids, which may play a role in metacyclogenesis, varied quantitatively among triatomine species in our study. Because the developmental success of a T. cruzi strain in its invertebrate host may depend on the triatomine species and the intestinal microbiota depends on the triatomine species, we suggest that a tripartite relationship is critical to the interactions between triatomines and T. cruzi. In addition, T. cruzi can also affect the microbiota composition upon triatomine infection, which, in turn, may affect the metabolic 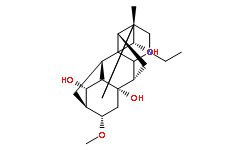 composition of triatomine feces, with significant feedback consequences on the parasite. Altogether, these observations suggest that a specific homeostatic system controlling the Estradiol Benzoate epidemiology of Chagas disease may exist. Therefore, it is necessary to determine how the three biological components of the triatomine-T. cruzi host-parasite interaction may affect the developmental success of the parasite. As such, this work is the first attempt to understand the complexity of the fecal metabolome in triatomines as a path for further investigations of triatomine vector competence for specific T. cruzi strains. Our work has opened many avenues for investigations concerning the role of individual metabolites in the interactions among T. cruzi, its triatomine vectors and the vertebrate host. Knowledge of these interactions is likely to generate new ways to understand the factors influencing parasite proliferation and spread as well as methods to control Chagas disease. The dosage compensation is an essential process that equalizes the expression levels of Xchromosomal genes between males and females. The complex is a model for studying chromosome-wide transcription regulation by histone hyperacetylation. It is actually known that the histone acetyltransferase MOF is tightly regulated by MSL1 and MSL3, and it even intervenes in the outcome of autophagy. In Drosophila, the complex also involves two functionally redundant non-coding RNAs ; in mammals, no associated RNA has been yet identified, and there are two MOF-MSL complexes. Both complexes are important for most of the histone 4 K16 acetylation in cells, for gene expression, cell-cycle regulation, DNA-damage repair and other regulatory processes in the nucleus. In addition to the MSL complex, MOF also resides in an NSL complex, that plays an important role in genome-wide chromatin modification and transcriptional regulation. The human 614-residue-long and the 1039-residue-long Drosophila MSL1s are predicted to contain no globular domains. Both, the scaffold of the MSL complex and dimerization of MSL1 coiled-coil region with the MSL2 protein are essential for male X-chromosome regulation in Drosophila.
composition of triatomine feces, with significant feedback consequences on the parasite. Altogether, these observations suggest that a specific homeostatic system controlling the Estradiol Benzoate epidemiology of Chagas disease may exist. Therefore, it is necessary to determine how the three biological components of the triatomine-T. cruzi host-parasite interaction may affect the developmental success of the parasite. As such, this work is the first attempt to understand the complexity of the fecal metabolome in triatomines as a path for further investigations of triatomine vector competence for specific T. cruzi strains. Our work has opened many avenues for investigations concerning the role of individual metabolites in the interactions among T. cruzi, its triatomine vectors and the vertebrate host. Knowledge of these interactions is likely to generate new ways to understand the factors influencing parasite proliferation and spread as well as methods to control Chagas disease. The dosage compensation is an essential process that equalizes the expression levels of Xchromosomal genes between males and females. The complex is a model for studying chromosome-wide transcription regulation by histone hyperacetylation. It is actually known that the histone acetyltransferase MOF is tightly regulated by MSL1 and MSL3, and it even intervenes in the outcome of autophagy. In Drosophila, the complex also involves two functionally redundant non-coding RNAs ; in mammals, no associated RNA has been yet identified, and there are two MOF-MSL complexes. Both complexes are important for most of the histone 4 K16 acetylation in cells, for gene expression, cell-cycle regulation, DNA-damage repair and other regulatory processes in the nucleus. In addition to the MSL complex, MOF also resides in an NSL complex, that plays an important role in genome-wide chromatin modification and transcriptional regulation. The human 614-residue-long and the 1039-residue-long Drosophila MSL1s are predicted to contain no globular domains. Both, the scaffold of the MSL complex and dimerization of MSL1 coiled-coil region with the MSL2 protein are essential for male X-chromosome regulation in Drosophila.
The design of time-series experiments to compare defense mutants with wild type for their PTI
Oxidative burst is critical for host-pathogen interactions. Many prior studies used methods involving dye staining, fluorescence, and/or chemiluminescence to detect ROS production. These methods are based on the activities of oxidase enzymes, such as peroxidase and NADPH oxidase, and therefore indirectly measure the levels of ROS in planta. Although providing relative 3,4,5-Trimethoxyphenylacetic acid quantitative information on ROS production, these methods do not offer precise subcellular localization of ROS, which could play a critical role in defense signal activation and transduction. The cerium chloride-based method, on the other hand, detects via TEM the electron dense precipitates resulting from the reaction of CeCl3 with H2O2, thus allowing a direct detection of H2O2 at the organelle levels. This method has been used in several plant-pathogen systems, such as Arabidopsis and lettuce-P. syringae, French bean�CXanthomonas, tomato/bean-Botrytis cinerea, legume-Rhizobia, tomato-nematode, and Solanum-Potato virus Y, and has provided spatial information of H2O2 accumulation in these plant pathosystems. We used this method for the first time to systematically examine when and where ROS is produced during PTI, ETS, and ETI. Our data have revealed temporal and spatial resolution of H2O2 localization during Arabidopsis-P. syringae interactions. We show that ETI is associated with much faster H2O2 accumulation than ETS, which is consistent with previous studies. Our data also show spatial difference in H2O2 accumulation, with ETI-induced H2O2 initially on the cell wall then inside of the cell and ETS-induced H2O2 beginning both inside and outside of the cell. H2O2 may contribute to cell death caused by DG34 and DG3 since the induction of H2O2 Mechlorethamine hydrochloride precedes the timing of massive cell death in the infected tissue. PTI-induced H2O2, on the other hand, accumulated much slowly and was only limited to the cell wall. Such temporal and spatial differences in H2O2 accumulation might reflect different mechanisms of plant resistance during PTI, ETS, and ETI. Pathogen-induced programmed cell death 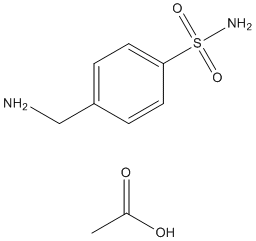 has been the focus in the studies of host-pathogen interactions. However, the change of host cell growth upon pathogen infection has been largely overlooked. Some animal pathogens are known to interfere with the host cell cycle machinery, activating cell growth and sometimes leading to tumorigenesis in animals. Pathogen-triggered cell growth has also been documented in some plants. For instance, nematodes induce host plants to form large cells, either resulting from fusion of the infected cells with its neighboring cells or from cell enlargement. The fungal pathogen powdery mildew was also shown to induce cell enlargement in Arabidopsis. These enlarged cells have increased nuclear DNA content, suggesting an activation of endoreplication. In addition, bacteria, such as Xanthomonas and Agrobacterium tumefaciens, can induce gall-like abnormal growths in their specific hosts, which could result from host cell enlargement and/or cell division. However, the mechanisms underlying host cell fate control upon pathogen infection have not been fully understood. Here we show that P. syringae also induces abnormal growths in Arabidopsis, which consist of enlarged mesophyll cells. The fact that the HrcC- strain induces more abnormal growths than both DG3 and DG34 strains suggests that PTI plays a major role in regulating cell growth. Consistent with this notion, our data further show that flg22- and perhaps also other PAMPs-induced PTI could act together to induce cell fate change in the host. Bacterial effectors were also reported to control host cell fate. Our data show that only the avirulent but not the virulent P. syringae strain induced minor abnormal.
has been the focus in the studies of host-pathogen interactions. However, the change of host cell growth upon pathogen infection has been largely overlooked. Some animal pathogens are known to interfere with the host cell cycle machinery, activating cell growth and sometimes leading to tumorigenesis in animals. Pathogen-triggered cell growth has also been documented in some plants. For instance, nematodes induce host plants to form large cells, either resulting from fusion of the infected cells with its neighboring cells or from cell enlargement. The fungal pathogen powdery mildew was also shown to induce cell enlargement in Arabidopsis. These enlarged cells have increased nuclear DNA content, suggesting an activation of endoreplication. In addition, bacteria, such as Xanthomonas and Agrobacterium tumefaciens, can induce gall-like abnormal growths in their specific hosts, which could result from host cell enlargement and/or cell division. However, the mechanisms underlying host cell fate control upon pathogen infection have not been fully understood. Here we show that P. syringae also induces abnormal growths in Arabidopsis, which consist of enlarged mesophyll cells. The fact that the HrcC- strain induces more abnormal growths than both DG3 and DG34 strains suggests that PTI plays a major role in regulating cell growth. Consistent with this notion, our data further show that flg22- and perhaps also other PAMPs-induced PTI could act together to induce cell fate change in the host. Bacterial effectors were also reported to control host cell fate. Our data show that only the avirulent but not the virulent P. syringae strain induced minor abnormal.
Affected remarkably by specific factors present in the biological compartments where aggregation occurs
The bacterial TDP-43 IBs characterized here were found to be highly toxic to cultured neuronal cells, particularly following their internalization in the cytoplasm, where they are at least in part ubiquitinated and phosphorylated. A significant toxicity was found using intracellularly delivered IBs where TDP-43 was present at a concentration as low as 1.7 mg/mL before internalisation. It is debated whether aggregation of TDP-43 in the cytoplasm of ALS and FTLD-U patients causes neurodegeneration due to formation of toxic protein aggregates or to the translocation of TDP-43 from the nucleus, which represents its physiological location, to the cytoplasm, or both. Our results show that delivery of exogenous TDP-43 into the cytoplasm occurs in the absence of a significant loss of endogenous TDP-43 in the nucleus. Indeed, the images acquired after delivery show cytoplasmic TDP-43 in the absence of detectable clearance of nuclear TDP-43. Thus, the non-amyloid, amorphous aggregates formed from TDP-43 are inherently highly toxic to neuronal cells, indicating that a gain of function mechanism caused by TDP-43 deposits is effective in such pathology. The data do not exclude that a loss of function mechanism originating from the cellular mistrafficking of TDP-43 also contributes to pathology, but shows the Pimozide inherent toxicity of TDP-43 aggregates. In conclusion we have shown, using bacterial IBs containing aggregated TDP-43 as a model system, that both FL and Ct TDP43 aggregates consist of non-amyloid assemblies that have an intrinsically high ability to cause neuronal dysfunction when delivered into the cytoplasm, contributing to elucidate the pathogenesis of TDP-43 proteinopathies such as FTLD-U and ALS. Enhanced TCR-driven calcium mobilization was observed in human LYPW620 carriers and in T cells from a mouse carrying a Diperodon knock-in R619W mutation in mouse Pep that is homologous to the human LYP R620W variation. Chang et al. identified a new dominant-negative isoform of LYP and proposed a model that reconciles “gain-of-function” and “loss-of-function” observations. Dai et al. recently reported a phenotype of enhanced TCR signaling and spontaneous autoimmunity in R619W knock-in mice. Analysis of the spectrum of phosphorylated molecules in TCR-stimulated Pep-R619W T cells suggested altered enzymatic specificity. In line with this  “altered function” model, a recent analysis of peripheral T cells from genotyped healthy subjects suggested that the LYP-R620W mutation can positively or negatively affect TCR signaling, depending on the biochemical readout assayed and on the stage of signaling. A prevailing model of thymocyte selection holds that TCR affinity for MHC/peptide ligand plays a central role in shaping the peripheral TCR repertoire. Deletion of autoreactive thymoyctes and agonist selection of regulatory T cells are two important mechanisms for establishing T cell tolerance that depend upon high-affinity interactions between TCR and ligand. Phenotyping of Ptpn22 knockout mice suggested that Pep might play a role in regulating thymic selection. Increased positive thymic selection has been reported in Ptpn22 KO mice and in two independently-generated Pep R619W knock-in mouse models, one of which developed spontaneous autoimmunity. Increased negative selection of H-Y transgenic male thymocytes mice was reported in one Pep R619W knock-in model, but thymic deletion was not altered in the other Pep R619W knock-in strain or in Ptpn22 KO mice. We reported increased TCR signaling in Ptpn22 KO thymocytes, correlating with increased thymic output of Treg in KO mice ; however, another group found no difference in thymic Treg percentages in an independently generated KO model.
“altered function” model, a recent analysis of peripheral T cells from genotyped healthy subjects suggested that the LYP-R620W mutation can positively or negatively affect TCR signaling, depending on the biochemical readout assayed and on the stage of signaling. A prevailing model of thymocyte selection holds that TCR affinity for MHC/peptide ligand plays a central role in shaping the peripheral TCR repertoire. Deletion of autoreactive thymoyctes and agonist selection of regulatory T cells are two important mechanisms for establishing T cell tolerance that depend upon high-affinity interactions between TCR and ligand. Phenotyping of Ptpn22 knockout mice suggested that Pep might play a role in regulating thymic selection. Increased positive thymic selection has been reported in Ptpn22 KO mice and in two independently-generated Pep R619W knock-in mouse models, one of which developed spontaneous autoimmunity. Increased negative selection of H-Y transgenic male thymocytes mice was reported in one Pep R619W knock-in model, but thymic deletion was not altered in the other Pep R619W knock-in strain or in Ptpn22 KO mice. We reported increased TCR signaling in Ptpn22 KO thymocytes, correlating with increased thymic output of Treg in KO mice ; however, another group found no difference in thymic Treg percentages in an independently generated KO model.