The Folinic acid calcium salt pentahydrate nuclear pore complex is a supramolecular protein assembly forming a highly selective channel embedded in the nuclear membrane. It regulates bidirectional nucleo-cytoplasmic transport for a large range of proteins and complexes too large to diffuse freely through the NPC. They are composed of numerous copies of,30 different nucleoporins, which have a well-assigned localisation, function and half-life, and are present as multiples of eight reflecting the highly conserved eight-fold axial symmetry of NPCs. The central substructure of the NPC is composed of transmembrane Nups that anchor the NPC to the nuclear envelope, scaffold Nups that constitute cornerstones during NPC biogenesis, and FG-Nups so-called because they contain extensive repeats of phenylalanine-glycine domains that form an unstructured mesh at the centre of the channel. Nup358/ RanBP2 and Nup214/CAN have been mapped exclusively to the cytoplasmic side of the NPC, where 50�C100 nm long flexible cytoplasmic filaments radiate from the NPC into the cytoplasm. Nup358/RanBP2 has been reported to be the major component of the cytoplasmic NPC filaments. Nup98 is a symmetrical nucleoporin, located on both the cytoplasmic and nuclear sides of the NPC. On the nuclear side of the NPC, Nups such as Nup153 and Nup98 associate with the nuclear basket and with the chromatin both in proximity of and away from the NPC. Many viruses depend on access to the nuclear compartment for replication and have evolved unique strategies to translocate into the nucleus. Retroviruses such as Murine Leukaemia Virus enter the nucleus during mitotic nuclear membrane disassembly, however other viruses such as herpesviruses and adenoviruses dock their capsids at the NPC and release their genome into the nucleus, while still others enter in the nucleus with their capsid. The Human Immunodeficiency Virus type 1, contrary to other orthoretroviruses, has evolved the ability to infect non-dividing cells through active nuclear Catharanthine sulfate import of its genome across the intact nuclear membrane through the NPC. Although several viral elements have been proposed to act as determinants of HIV-1 nuclear import, most notably integrase and the central DNA Flap, it is commonly accepted that HIV-1 depends on host cell proteins to achieve translocation. However, increasing data supports the presence of HIV-1 capsid at the nuclear pore and/or acting as a determinant of HIV-1 nuclear import. Uncoating certainly also occurs during cytoplasmic transport, and possibly accounts for the majority of incoming viral complexes. However, these may correspond to viral complexes destined for or undergoing degradation, for instance following entry by endocytosis. Our work shows that HIV-1 capsid interacts with Nup358/RanBP2 and that depletion of Nup358/RanBP2 impairs arrival of HIV-1 complexes at the nuclear envelope, thus confirming the presence of HIV-1 capsid cores at the nuclear membrane. However, our study does not show any effect for Nup358 in integration, since the strong reduction in proviral integration is simply due to a strong nuclear import defect and considering the exclusive cytoplasmic location of Nup358/RanBP2 we do not expect to find its potential viral partner in the nucleus. We cannot exclude that the absence of this nucleoporin could affect HIV-1 site integration but that may reflect 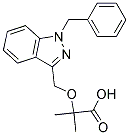 the change of chromatin environment associated with depletion of Nup358. In this work, we identified Nup358/RanBP2 as docking factor for HIV-1 capsid using independent techniques to assess docking and interaction. We are the first to identify the binding of Nup358/ RanBP2 to HIV-1 in vitro assembled CA-NC complexes suggesting that this interaction is important for HIV-1 nuclear import. Nup358/RanBP2 is one of over 20 human proteins that contain a cyclophilin-like domain.
the change of chromatin environment associated with depletion of Nup358. In this work, we identified Nup358/RanBP2 as docking factor for HIV-1 capsid using independent techniques to assess docking and interaction. We are the first to identify the binding of Nup358/ RanBP2 to HIV-1 in vitro assembled CA-NC complexes suggesting that this interaction is important for HIV-1 nuclear import. Nup358/RanBP2 is one of over 20 human proteins that contain a cyclophilin-like domain.
Month: June 2019
Clinically relevant tumor samples are therefore likely to represent the cumulative result of undirected generation of variance followed
Participate in the network transformations associated with tumorigenesis. A high level view of network rearrangements can be seen by comparing modules in AN and TU. Many significant overlaps were seen indicating that the two tissues were far from randomly organized with respect to each other. Closer examination revealed support for the disruption and creation of co-expression networks found with gene-gene differential correlation data. For example, the genes in the largest module in AN significantly overlapped the genes in eight different TU modules as well as genes that could not be placed in the TU network, consistent with disruption of the AN-turquoise module. Similarly, genes in the largest module in TU, overlapped genes in 10 AN modules as well as genes in AN-grey, consistent with ![]() the creation of TU-turquoise as part of HCC tumorigenesis. Consistent with the above interpretation of the AN and TU module overlaps was the finding that the differentially correlated genes were enriched in many of the modules. For example, 22 of the 25 AN modules and 13 of the 20 TU modules were enriched for genes containing differentially correlated genes, indicating that a majority of the subnetworks in the AN tissue representing many different biological functions were disrupted as a result of the formation and progression of the tumor, resulting in a higher level re-organization. To assess possible biological functions represented by the networks, each module was tested for over-representation of genes from individual gene ontology categories. Given the large scale reorganizations between AN and TU, GO terms that were most significantly enriched for each module compared to all other modules were defined. The purpose of this was to begin to define biological pathways that were uniquely disrupted in AN tissue and uniquely Orbifloxacin created in TU tissue. Examples of this are the findings that components of the ribosome were uniquely enriched in the TU-grey60 module, and aspects of macrophage function were uniquely enriched in the AN-coral module. In both cases these terms were not enriched in any other module of either tissue. Many unique or relative enrichments of GO terms were found for 14 of the 25 AN modules and 12 of the 21 TU modules representing a broad array of biological functions coherently represented in the networks and altered between tissues in HCC tumorigenesis. Ribosome components have been clearly implicated in tumor initiation and progression in numerous cancer types. In particular it has been argued that altered translation Cinoxacin facilitates expression of many proliferation associated genes and may also regulate the endothelial to mesenchymal transition which is thought to be important in invasion and metastasis. The enrichment of TU-survival genes in TU-grey60 and TU-lightyellow suggests that translation and ribosome biogenesis were selected for alteration during tumorigenesis, but remained rate limiting factors in progression. Copy number aberrations are widely observed in solid tumors and are likely the result of altered fidelity of DNA replication, repair, checkpoints and/or chromosome segregation. These processes leading to sCNV are by their nature intrinsic features of cancer cells and occur in an undirected manner in terms of chromosome location and direction of change. Since generation of sCNV is ongoing and will lead to neutral, increased, or decreased fitness of the corresponding cell in its environment, different derivatives will have consequently different abilities to grow and survive, therefore leading to evolution of the tumor over time.
the creation of TU-turquoise as part of HCC tumorigenesis. Consistent with the above interpretation of the AN and TU module overlaps was the finding that the differentially correlated genes were enriched in many of the modules. For example, 22 of the 25 AN modules and 13 of the 20 TU modules were enriched for genes containing differentially correlated genes, indicating that a majority of the subnetworks in the AN tissue representing many different biological functions were disrupted as a result of the formation and progression of the tumor, resulting in a higher level re-organization. To assess possible biological functions represented by the networks, each module was tested for over-representation of genes from individual gene ontology categories. Given the large scale reorganizations between AN and TU, GO terms that were most significantly enriched for each module compared to all other modules were defined. The purpose of this was to begin to define biological pathways that were uniquely disrupted in AN tissue and uniquely Orbifloxacin created in TU tissue. Examples of this are the findings that components of the ribosome were uniquely enriched in the TU-grey60 module, and aspects of macrophage function were uniquely enriched in the AN-coral module. In both cases these terms were not enriched in any other module of either tissue. Many unique or relative enrichments of GO terms were found for 14 of the 25 AN modules and 12 of the 21 TU modules representing a broad array of biological functions coherently represented in the networks and altered between tissues in HCC tumorigenesis. Ribosome components have been clearly implicated in tumor initiation and progression in numerous cancer types. In particular it has been argued that altered translation Cinoxacin facilitates expression of many proliferation associated genes and may also regulate the endothelial to mesenchymal transition which is thought to be important in invasion and metastasis. The enrichment of TU-survival genes in TU-grey60 and TU-lightyellow suggests that translation and ribosome biogenesis were selected for alteration during tumorigenesis, but remained rate limiting factors in progression. Copy number aberrations are widely observed in solid tumors and are likely the result of altered fidelity of DNA replication, repair, checkpoints and/or chromosome segregation. These processes leading to sCNV are by their nature intrinsic features of cancer cells and occur in an undirected manner in terms of chromosome location and direction of change. Since generation of sCNV is ongoing and will lead to neutral, increased, or decreased fitness of the corresponding cell in its environment, different derivatives will have consequently different abilities to grow and survive, therefore leading to evolution of the tumor over time.
The ability of the recruited cells characterized by tumor suppressor loss to contribute to high-grade glioma structures
We performed transplantation experiments using the bacTRAP system. Transplanted gliomas induced by hPDGFb-driven glioma cells in hosts with altered tumor suppressor function showed higher percentages of the overall recruitment than in the wild-type hosts, indicating that complete or partial tumor suppressor loss may enhance the ability of the cells to be  recruited. Although gliomas induced by transplantation of non-fluorescent Pten-deleted Ras-driven murine glioma cells showed recruitment of proliferating host brain cells into the glioma mass, the overall amount of recruited cells into Ras-driven tumors and the number of tumors that showed large regions of recruitment was significantly less than that seen with the hPDGFb-induced gliomas. An unbiased way to define a cell population lies in Gomisin-D identification of its gene expression signature and subsequent comparison to gene expression signatures of known normal or cancer cells to define its position on the axis of tumorigenesis. To quantify similarities and differences in the expression profiles of recruited cells and tumor cells using microarray analysis, we used the bacTRAP technology that allows immunoprecipitation of polysomes from specific cell types in vivo. RCAS/tv-a system allows to closely model the biology and histopathology of human oligodendrogliomas; although we have not performed direct comparisons between our mouse model and human glioma classifications, in many ways, hPDGFb-driven murine gliomas may mimic the proneural subclass of human GBMs. Our data suggests that olig2-expressing cells recruited in PDGF-induced murine gliomas can have similar morphologic, proliferative and functional characteristics as olig2expressing tumor cells derived from the cell-of-origin, with the caveat that a part of the similarity between the recruited and tumor olig2 cells may stem from their proliferative behavior. Extensive similarities between polysome-associated transcriptome of recruited versus glioma olig2 cells call for further evaluation of the precise definitions and criteria applied to terms “tumor” and “normal”, and indicate similar gene expression character of these cells. Furthermore, tumor suppressor loss and acquisition of mutations typically found in human gliomas allow these murine recruited cells to occupy large areas of the tumor and at times become the predominant cellular component of the glioma mass, completely independent of the cell-of-origin. Thus, the clonal expansion that overtakes glioma bulk during tumor progression need not be derived from glioma cell-of-origin. Transformation is the process of self-autonomous acquisition of the sufficient oncogenic alterations that change a normal cell into the tumor cell. Unlike epithelial cancers, the “normal” or stromal component of a glioma is composed of cells of the same lineage as the tumor cells. The inherent implication of this observation is that extracellular factors that promote glioma tumor cell growth may similarly affect glioma stroma. In our glioma model, initially normal recruited cells not derived from the glioma-initiating cellof-origin can be stimulated by hyperproduction of a growth factor receptor ligand, induced to proliferate, and driven to acquire various genetic aberrations and Cinoxacin aberrant expression profiles as gliomas progress, presumably because of the tumor microenvironment created by the progeny of the cell-of-origin. The process of acquisition and selection for such genetic and/or epigenetic alterations due to the pressures of the glioma microenvironment may be better termed “corruption”.
recruited. Although gliomas induced by transplantation of non-fluorescent Pten-deleted Ras-driven murine glioma cells showed recruitment of proliferating host brain cells into the glioma mass, the overall amount of recruited cells into Ras-driven tumors and the number of tumors that showed large regions of recruitment was significantly less than that seen with the hPDGFb-induced gliomas. An unbiased way to define a cell population lies in Gomisin-D identification of its gene expression signature and subsequent comparison to gene expression signatures of known normal or cancer cells to define its position on the axis of tumorigenesis. To quantify similarities and differences in the expression profiles of recruited cells and tumor cells using microarray analysis, we used the bacTRAP technology that allows immunoprecipitation of polysomes from specific cell types in vivo. RCAS/tv-a system allows to closely model the biology and histopathology of human oligodendrogliomas; although we have not performed direct comparisons between our mouse model and human glioma classifications, in many ways, hPDGFb-driven murine gliomas may mimic the proneural subclass of human GBMs. Our data suggests that olig2-expressing cells recruited in PDGF-induced murine gliomas can have similar morphologic, proliferative and functional characteristics as olig2expressing tumor cells derived from the cell-of-origin, with the caveat that a part of the similarity between the recruited and tumor olig2 cells may stem from their proliferative behavior. Extensive similarities between polysome-associated transcriptome of recruited versus glioma olig2 cells call for further evaluation of the precise definitions and criteria applied to terms “tumor” and “normal”, and indicate similar gene expression character of these cells. Furthermore, tumor suppressor loss and acquisition of mutations typically found in human gliomas allow these murine recruited cells to occupy large areas of the tumor and at times become the predominant cellular component of the glioma mass, completely independent of the cell-of-origin. Thus, the clonal expansion that overtakes glioma bulk during tumor progression need not be derived from glioma cell-of-origin. Transformation is the process of self-autonomous acquisition of the sufficient oncogenic alterations that change a normal cell into the tumor cell. Unlike epithelial cancers, the “normal” or stromal component of a glioma is composed of cells of the same lineage as the tumor cells. The inherent implication of this observation is that extracellular factors that promote glioma tumor cell growth may similarly affect glioma stroma. In our glioma model, initially normal recruited cells not derived from the glioma-initiating cellof-origin can be stimulated by hyperproduction of a growth factor receptor ligand, induced to proliferate, and driven to acquire various genetic aberrations and Cinoxacin aberrant expression profiles as gliomas progress, presumably because of the tumor microenvironment created by the progeny of the cell-of-origin. The process of acquisition and selection for such genetic and/or epigenetic alterations due to the pressures of the glioma microenvironment may be better termed “corruption”.
This study aimed to determine array of various diseases and pathophysiological conditions
Intrauterine growth restriction is associated with increased perinatal morbidity and mortality and with increased risk of adult diseases such as diabetes, hypertension, and coronary artery disease. Impaired growth may persist during childhood despite optimum nutrition. Although a poorly growing fetus can be relatively easily identified by obstetric ultrasound, the therapeutic options are limited. Thus, sustained poor growth in utero frequently results in the fetus being delivered, with the attendant morbidity and mortality of preterm birth. Further, preterm birth is itself associated with hypertension, diabetes and insulin resistance, ischemic heart disease and stroke in later life. It is unclear whether intervention early in postnatal life can ameliorate these increased risks. For example, accelerated postnatal growth may increase the long-term risks associated with reduced size at birth. Therefore, attempting to reverse IUGR in utero may represent the optimum approach. In developed nations, IUGR typically is caused by placental insufficiency, resulting in a reduced fetal nutrient supply. Thus, clinical attempts to improve fetal growth by maternal supplementation with protein or oxygen were unsuccessful. Attempts to increase placental blood flow with sildenafil citrate in IUGR ovine pregnancies resulted in adverse outcome, and supplementation of pregnant women carrying severely IUGR fetuses with L-arginine had no effect on fetal growth. Therefore, a treatment that bypasses the placenta may provide the most promising approach. Insulin-like 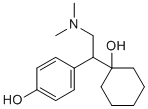 growth factor-1 is the primary endocrine regulator of fetal growth in late gestation. Birth weight is positively associated with IGF-1 concentrations in umbilical cord blood, and circulating fetal IGF-1 concentrations are lower in IUGR pregnancies. Acute high-dose intravenous IGF-1 infusion has anabolic effects on fetal sheep, stimulating substrate uptake and inhibiting protein breakdown. However, while continuous vascular access to the fetus is not practicable in IUGR treatment, the amniotic fluid compartment is routinely accessed in clinical practice. IGF-1 administered into the amniotic fluid in sheep is swallowed and taken up across the fetal gut to circulate Albaspidin-AA intact in the fetus. Further, Ginsenoside-Ro administration of low dose intra-amniotic IGF-1 thrice weekly increases growth of ovine fetuses with IUGR induced by placental embolization. However, for intraamniotic IGF-1 treatment to be clinically useful, a less frequent administration is required. In addition, circulating fetal concentrations of IGF-1 following treatment were either unaltered or actually decreased, with down-regulation of fetal hepatic igf1 mRNA levels. These data suggest that the increased fetal growth after intra-amniotic IGF-1 is unlikely to be mediated directly by circulating IGF-1. One possible mechanism is via effects on placental nutrient transport. Glucose is transported across the placenta by facilitated diffusion, mediated by glucose transporters. SLC2A1 and SLC2A3 are the major glucose transporter isoforms in the placenta of ruminants and rodents, with SLC2A4 also described in humans and rats. In contrast, amino acids are transported across the placenta by active transport mediated by numerous different amino acid transporters, many of which have several isoforms. Recent studies have suggested that changes in placental amino acid transport, possibly mediated by altered activity of mammalian target of rapamycin, precede growth restriction and may therefore directly contribute to decreased fetal growth. IGF-1 is known to alter activity and/or expression of nutrient transporters in different placental preparations.
growth factor-1 is the primary endocrine regulator of fetal growth in late gestation. Birth weight is positively associated with IGF-1 concentrations in umbilical cord blood, and circulating fetal IGF-1 concentrations are lower in IUGR pregnancies. Acute high-dose intravenous IGF-1 infusion has anabolic effects on fetal sheep, stimulating substrate uptake and inhibiting protein breakdown. However, while continuous vascular access to the fetus is not practicable in IUGR treatment, the amniotic fluid compartment is routinely accessed in clinical practice. IGF-1 administered into the amniotic fluid in sheep is swallowed and taken up across the fetal gut to circulate Albaspidin-AA intact in the fetus. Further, Ginsenoside-Ro administration of low dose intra-amniotic IGF-1 thrice weekly increases growth of ovine fetuses with IUGR induced by placental embolization. However, for intraamniotic IGF-1 treatment to be clinically useful, a less frequent administration is required. In addition, circulating fetal concentrations of IGF-1 following treatment were either unaltered or actually decreased, with down-regulation of fetal hepatic igf1 mRNA levels. These data suggest that the increased fetal growth after intra-amniotic IGF-1 is unlikely to be mediated directly by circulating IGF-1. One possible mechanism is via effects on placental nutrient transport. Glucose is transported across the placenta by facilitated diffusion, mediated by glucose transporters. SLC2A1 and SLC2A3 are the major glucose transporter isoforms in the placenta of ruminants and rodents, with SLC2A4 also described in humans and rats. In contrast, amino acids are transported across the placenta by active transport mediated by numerous different amino acid transporters, many of which have several isoforms. Recent studies have suggested that changes in placental amino acid transport, possibly mediated by altered activity of mammalian target of rapamycin, precede growth restriction and may therefore directly contribute to decreased fetal growth. IGF-1 is known to alter activity and/or expression of nutrient transporters in different placental preparations.
The nature and fate of these cells not derived from the glioma cell-of-origin has not be gliomas
Show expression patterns that are correlated with PDGFR signaling, a pattern with prominent expression of OLIG2 and other genes involved in CNS development referred to as “proneural”. PDGF ligands are upregulated in at least a third of surgical glioma samples and human glioma cell lines. The importance of PDGF signaling is underscored in genetically engineered rodent gliomas, where overproduction of human PDGFb ligand is sufficient to induce gliomagenesis in a dosedependent manner and allows to recapitulate the histologic, etiologic and pathobiologic character of the PDGF subset of human gliomas. Additionally, infusion of PDGF into the ventricles induces proliferation of the SVZ, resulting in lesions with some characteristics of gliomas. Similar to human gliomas, mouse gliomas are cellularly and molecularly heterogeneous. Glioma progression in humans is associated with deletion of the INK4A/ARF locus and loss of PTEN expression resulting in activation of Akt. The standard view of gliomagenesis is that sequential mutations occur and accumulate in cells derived from the glioma cell-of-origin. Indeed, many surgical GBM samples in patients appear clonal, with all tumor cells seemingly derived from the same cell; however, this may not necessarily mean they are derived from the cell-of-origin. Cellular heterogeneity and reports of human gliomas comprised of several genetically unrelated clones suggest the possibility of oncogenic transformation in cells not derived from the glioma cell-of-origin. The interconversion between human glioma subtypes upon recurrence and the existence of recurrent gliomas that lack mutations or deletions found in the original tumor further indicate the possibility for an expansion of an aggressive clone not arising from the cell-of-origin. In fact, PDGF-induced gliomas arising in both adult and neonatal rats have been shown to contain normal stem and progenitor cells “recruited” into glioma mass and induced to proliferate, indicating that proliferative stem-like portions of the tumor can arise from normal progenitors. However, the Mepiroxol precise nature and specific functional characteristics of these “recruited” stem or progenitor cells have not been described. Genetic analysis of surgical samples of human gliomas merely provides retrospective static information with regards to tumor evolution; lineage tracing from the cell-of-origin cannot be done in humans. Moreover, identifying and distinguishing GBM cells from the surrounding stroma is not a trivial task – glioma cells are often defined histologically, demonstrating high mitotic indices, expression of stem or progenitor cell markers, abnormal global gene expression patterns, presence of genetic alterations, and the ability to serially transplant the disease. To investigate cellular contributions and structural/functional characteristics of “recruited” cells in murine gliomas during tumor progression, we used RCAS/tv-a and the Gomisin-D bacTRAP systems. Defining tumor cells by histologic criteria, genetic analysis, global gene expression profiling and transplantation studies, we studied the clonality of mouse gliomas with respect to the cell-of-origin. Here we show that in murine gliomas induced by human PDGFb, glioma progression can occur 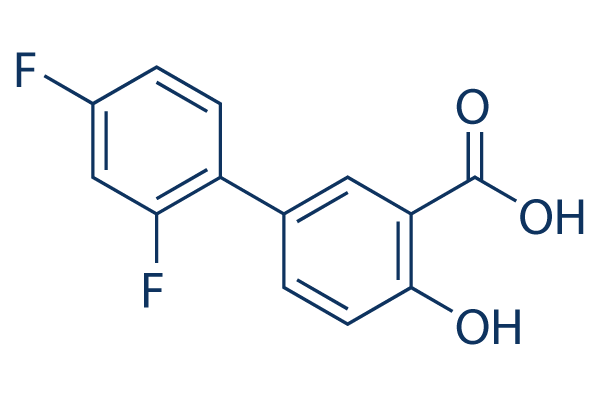 by expansion of the recruited cells, and that these cells unrelated to glioma cell-of-origin can be corrupted to become bona fide tumor. It has been recently shown that gliomas induced in adult or neonatal rats by hPDGFb-expressing retroviruses contain stem or progenitor-like cells expressing neural markers, that are contributing to glioma mass and are induced to proliferate by glioma environment.
by expansion of the recruited cells, and that these cells unrelated to glioma cell-of-origin can be corrupted to become bona fide tumor. It has been recently shown that gliomas induced in adult or neonatal rats by hPDGFb-expressing retroviruses contain stem or progenitor-like cells expressing neural markers, that are contributing to glioma mass and are induced to proliferate by glioma environment.